Lecture 04
第4讲学习讲义:DNA, Chromosomes and Genome


对应课件:Lecture_4_DNA_Chromosome_and_Genome.pdf
这一讲的关键词是:包装、组织、可读性、遗传性。
很多初学者会觉得 DNA 只是一条双螺旋,但真正的细胞里,DNA 从来不是裸露存在的。它必须被压缩、组织、标记,还要在需要时重新打开。这就是染色体和染色质研究的核心。
这讲的核心问题
- DNA 双螺旋有哪些最基础的结构特征?
- 为什么 2 米长的人类 DNA 能装进细胞核?
- 染色质如何既被压缩又保留可读取性?
- 染色质状态如何参与基因调控和表观遗传?
一、先把 DNA 双螺旋的基本事实讲清楚
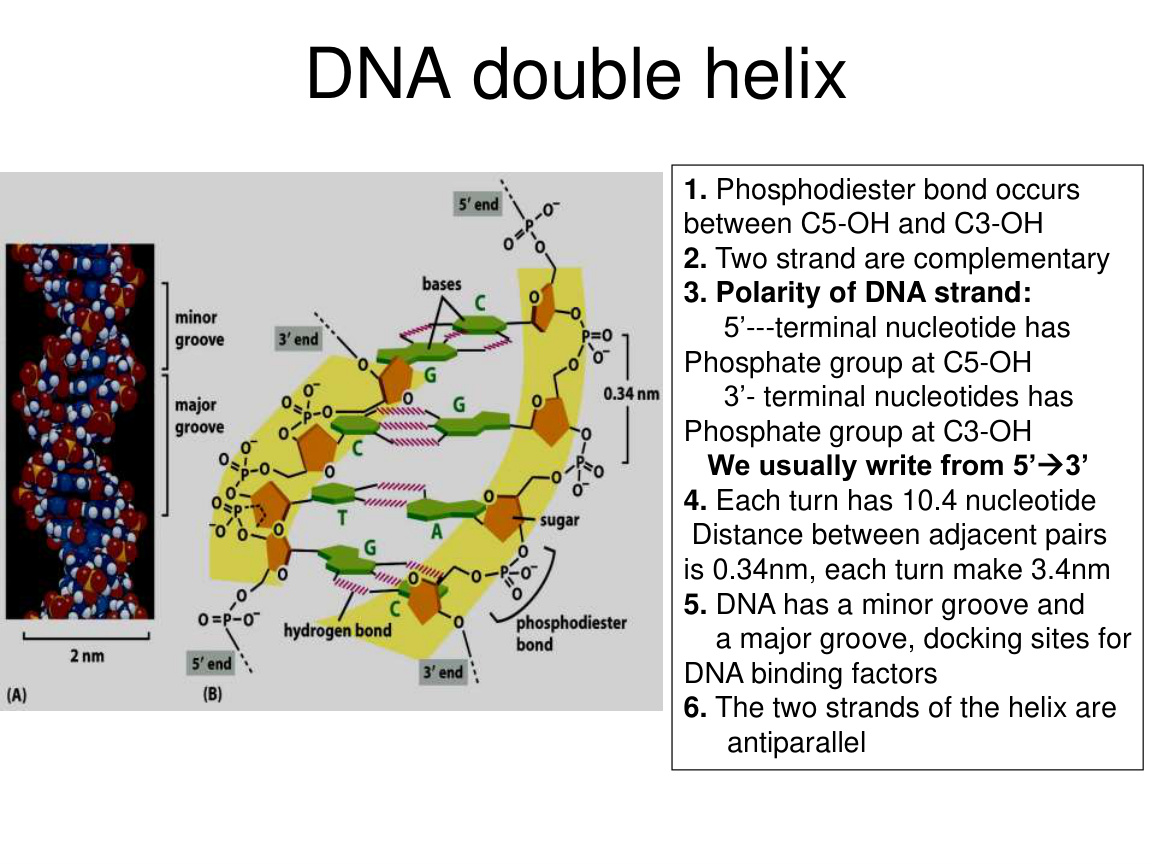
1. DNA 的化学骨架
DNA 链中核苷酸通过磷酸二酯键连接。
这使 DNA 具有:
- 稳定骨架。
- 方向性。
2. DNA 两条链互补
互补配对意味着:
- 一条链的信息可以决定另一条链。
- 复制和修复都有了模板基础。
3. DNA 有方向性
每条链都有 5' 端和 3' 端。
生物学里默认把序列写成 5' -> 3'。
这个方向性不是记号游戏,而是决定:
- 聚合酶合成方向。
- 启动子和基因方向。
- 复制前导链与后随链差异。
4. 两条链是反向平行 antiparallel
一条是 5' -> 3',另一条是 3' -> 5'。
这直接决定复制和转录时酶怎么读取模板。
5. 大沟和小沟
DNA 双螺旋有 major groove 和 minor groove。
这很重要,因为很多 DNA 结合蛋白并不是把 DNA 拆开来读,而是:
- 从沟槽表面的化学特征识别序列。
二、染色体、染色质、基因组、核型:四个基础概念一定要分开
1. Chromatin 染色质
DNA 和蛋白质的复合物,尤其包括组蛋白。
2. Chromosome 染色体
一条长 DNA 分子与相关蛋白打包形成的结构,在有丝分裂时最容易在显微镜下看到。
3. Genome 基因组
一个细胞所含全部遗传信息的总和。
4. Karyotype 核型
把分裂中期染色体按大小和形态排列展示出来的图。
关系可以这样记:
- 基因组是全部信息。
- 染色体是信息被分配到的独立包装单元。
- 染色质是 DNA 在细胞中的实际物质状态。
- 核型是观察染色体的展示方式。
三、人类基因组为什么是一个包装奇迹
课件强调:
- 人类有 23 对染色体。
- 总 DNA 长度大约可达 2 米。
- 但却装在一个很小的细胞核里。
这提出了一个核心问题:
- DNA 必须极度压缩。
- 但又不能压缩到完全失去功能。
因此染色质不是单纯“卷起来”,而是有层次、有动态、有调控地组织。
四、染色体的基本构造:着丝粒、端粒、复制后的姐妹染色单体
1. 着丝粒 centromere
着丝粒是染色体分离时的关键区域。
它的核心作用是:
- 组装动粒。
- 让纺锤丝附着。
- 保证姐妹染色单体被正确拉开。
2. 端粒 telomere
端粒位于线性染色体末端。
功能包括:
- 保护染色体末端不被误认为断裂。
- 缓冲末端复制问题。
3. 复制后的染色体状态
DNA 复制之后,一条染色体会形成两条姐妹染色单体,后面在分裂时再分开。
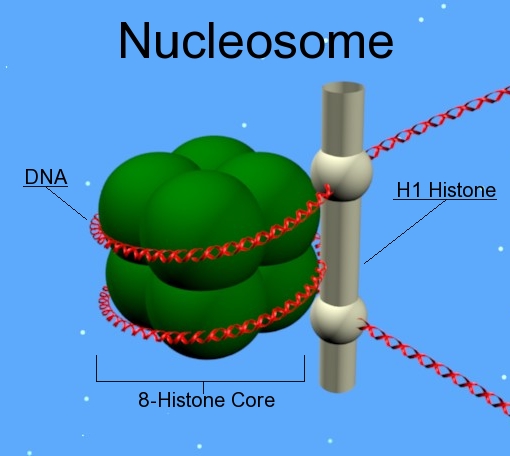
五、核小体:真核染色质的基本单位
1. 什么是核小体 nucleosome
核小体是 DNA 缠绕在组蛋白八聚体外形成的基本包装单位。
你可以把它想成:
- DNA 是线。
- 组蛋白核心像线轴。
- DNA 缠在线轴上。
2. 组蛋白八聚体 histone octamer
核心通常包括:
- H2A、H2B、H3、H4 各两份。
H1 通常不在核心八聚体内,但对更高层级压缩很重要。
3. 为什么核小体重要
它同时解决两件事:
- 帮助 DNA 压缩。
- 提供调控平台。
因为 DNA 缠到组蛋白上以后:
- 有些区域更难被蛋白访问。
- 有些区域在重塑后又能暴露出来。
六、核小体不是静止的,而是动态的
这特别重要,因为如果核小体完全固定,DNA 上很多基因根本读不出来。
动态性体现为:
- DNA 可在组蛋白表面短暂松开。
- 组蛋白可被替换。
- 核小体位置可移动。
- 核小体可被部分或整体移除。
七、ATP 依赖的染色质重塑
这类复合物使用 ATP 提供能量,去改变核小体状态,例如:
- 滑动核小体位置。
- 移除核小体。
- 重组核小体。
其生物学意义是:
- 让本来被包裹的 DNA 暴露出来。
- 或把 DNA 重新封闭起来。
因此,染色质调控本质上是“信息可访问性”的调控。
八、从核小体到高级压缩:DNA 如何层层打包
你现在不必死记每一种历史模型,但要理解层次感:
- DNA 双螺旋。
- 核小体串珠样结构。
- 更高级纤维化压缩。
- 环状结构 loop。
- 大尺度染色体区域。
- 有丝分裂期最致密染色体。
H1 的作用
你可以把 H1 理解成:
- 有助于稳定核小体间连接和更高层折叠的辅助组蛋白。
九、染色体在细胞核内不是乱塞的:有空间组织
细胞核不是“DNA 和蛋白的均匀汤”,而是高度分区的空间系统。
1. 染色体领地 chromosome territories
不同染色体在细胞核中往往占据相对偏好的区域,而不是完全随机混合。
2. 染色质环 loop
某些基因调控依赖远距离 DNA 片段靠近。
环状结构可以让:
- 增强子接近启动子。
- 特定调控区协同工作。
3. 核内功能区室
例如:
- Nucleolus 核仁。
- Splicing speckles 剪接斑。
- DNA damage foci DNA 损伤灶。
这些都说明:
- 空间组织本身就是调控的一部分。
十、常染色质与异染色质:打开和关闭的两种典型状态
1. Euchromatin 常染色质
一般特点:
- 相对松散。
- 转录更活跃。
- 更容易被访问。
2. Heterochromatin 异染色质
一般特点:
- 更紧密。
- 转录活性较低。
- 常与重复序列、结构区域相关。
但要注意:
- 它们不是绝对的二元开关。
- 更像一条连续谱,两端是开放和紧闭的典型状态。
十一、染色质调控与表观遗传:遗传的不只是 DNA 序列
1. 遗传 inheritance 不只有序列
传统遗传关注:
- 碱基序列如何代代相传。
表观遗传关注:
- 在不改变 DNA 序列的前提下,某些功能状态如何被维持和传递。
2. 组蛋白修饰 histone modifications
常见类型包括:
- 乙酰化。
- 甲基化。
- 磷酸化。
- 泛素化。
这些修饰会:
- 改变组蛋白和 DNA 的相互作用。
- 招募特定 reader 蛋白。
- 改变染色质开放程度。
3. Histone code 组蛋白密码
这个概念的意思不是存在一套像遗传密码那样固定的一一对应表,而是:
- 不同修饰组合携带不同调控含义。
- 这些组合可被蛋白识别,并转化成功能结果。
十二、读者、写者、擦除者与重塑复合物
这套思路很重要:
- Writer:添加修饰。
- Eraser:去除修饰。
- Reader:识别修饰并执行后续作用。
- Remodeler:改变核小体组织状态。
这说明染色质调控不是单一事件,而是一个会自我强化、也会被阻断的网络。
十三、组蛋白变体 histone variants
含义是:
- 有些位置并不使用“标准组蛋白”,而是替换成特殊版本。
这些变体可以改变:
- 核小体稳定性。
- 染色体特定区域身份。
- 特定 DNA 过程的效率。
十四、边界与位置效应:染色质状态会传播
这说明染色质状态有扩散趋势:
- 一个沉默区域可能向附近扩展。
- 一个活跃区域也可能影响相邻区域。
边界元件的作用是:
- 阻止某种状态无限扩散。
位置效应 position effect 则说明:
- 同一个基因,放在基因组不同位置,表达可能不同。
十五、染色质状态如何被继承
这意味着在细胞分裂后:
- 新细胞并不是从零开始决定所有染色质状态。
- 原有组蛋白修饰、核小体分布、相关调控蛋白会帮助“重建”原状态。
这正是表观遗传记忆的核心。
十六、把这一讲串成一句话
DNA 双螺旋只是遗传信息的基础形式;在真核细胞中,DNA 被组蛋白包装成动态的核小体和更高层染色质结构;这些结构不仅压缩 DNA,更决定 DNA 是否可被访问,并通过组蛋白修饰、变体、重塑和空间组织参与基因调控与表观遗传继承。
十七、常见误区
- “DNA 包装就是为了省空间”不完整。包装也直接参与调控。
- “核小体把 DNA 挡住了,所以一定抑制表达”不完全对。关键是动态重塑和局部状态。
- “表观遗传就是 DNA 甲基化”太窄。组蛋白修饰、核小体结构和空间组织也很重要。
- “染色体在细胞核里是随机漂浮的”通常不对,存在明显空间组织规律。
十八、你必须会的关键词
- Chromatin:染色质。
- Chromosome:染色体。
- Genome:基因组。
- Karyotype:核型。
- Nucleosome:核小体。
- Histone octamer:组蛋白八聚体。
- Euchromatin:常染色质。
- Heterochromatin:异染色质。
- Histone modification:组蛋白修饰。
- Chromatin remodeling:染色质重塑。
- Histone variant:组蛋白变体。
- Epigenetic inheritance:表观遗传继承。
十九、自测题
1. 核小体为什么是染色质的基本单位?
答题关键:
- 是 DNA 与组蛋白形成的基本包装模块。
- 同时承担压缩与调控平台功能。
2. 为什么说染色质不是静态结构?
答题关键:
- 核小体可移动、替换、重塑。
- DNA 可在局部暴露或封闭。
3. 常染色质和异染色质的典型差异是什么?
答题关键:
- 开放程度、转录活性、可访问性不同。
4. 什么是表观遗传继承?
答题关键:
- DNA 序列不变,但染色质状态和表达倾向可被传递。
二十、考前速记版
- DNA 有方向性、互补性和大沟小沟。
- 染色质是 DNA 与蛋白的复合物。
- 核小体是真核 DNA 包装基本单位。
- 染色质包装是动态且可调控的。
- 常染色质更开放,异染色质更压缩。
- 组蛋白修饰、变体和重塑复合物共同决定染色质状态。
- 染色质状态可参与表观遗传记忆。
二十一、深入扩展:核小体为什么既是“包装单位”又是“调控单位”
如果核小体只是为了把 DNA 缠起来省空间,那它更像被动包装材料。但真实情况是,它同时决定哪些 DNA 片段更容易被访问。
为什么核小体位置很重要
因为只要一个转录因子结合位点刚好被核小体遮住:
- 对应蛋白就不容易接近它。
- 该位点的功能就会被抑制。
反过来,如果核小体移动后把位点暴露:
- 该区域就更可能被激活。
所以“核小体放在哪里”本身就是调控信息。
二十二、为什么组蛋白尾巴是修饰热点
组蛋白尾巴通常比较伸展,暴露在核小体外侧,更适合作为:
- 化学修饰位点。
- reader 蛋白识别平台。
这使它们像核小体上的“控制面板”。不同修饰组合能改变:
- 组蛋白和 DNA 的相互作用强弱。
- 周围蛋白是否被招募。
- 局部染色质更偏开放还是关闭。
二十三、为什么不能把 histone code 理解成死板密码本
更好的理解是:
- 它像语境相关的组合信号系统。
- 同一个修饰在不同位置、不同时间、不同组合里,意义可能不同。
所以“组蛋白密码”不是一套固定查表题,而是一个需要结合上下文解释的调控语言。
二十四、三维基因组为什么越来越重要
以前很多人只从线性 DNA 序列理解基因调控,但实际上:
- 远距离增强子能不能接触到启动子,取决于三维折叠。
- 某个区域靠近核膜还是靠近核内活跃区,会影响表达倾向。
因此基因组不仅有“序列信息”,还有“空间组织信息”。
二十五、如果你想真正理解表观遗传,最简单的抓手是什么
先抓住这句:
- 相同 DNA 序列可以处于不同功能状态,而且这种状态可以在细胞分裂后延续。
这就是表观遗传最核心的直觉。
后面你再把它具体化成:
- 组蛋白修饰。
- 核小体分布。
- 染色质开放程度。
- DNA 甲基化。
- 空间组织。
这样理解会比一上来背名词更稳。