Lecture 09
第9讲学习讲义:细胞膜结构与功能

对应课件:lecture09_Membrane_Structure_Functions.pdf
这一讲的主线是:细胞膜不是一层被动的包裹膜,而是由脂质、蛋白质和糖链共同构成的动态功能平台。 它既定义细胞和细胞器的边界,也负责选择性通透、信号感知、能量转化、细胞识别和细胞骨架连接。
这讲的核心问题
- 为什么脂质能自发形成双分子层?
- 真核细胞膜主要由哪些脂质组成?
- 质膜两侧为什么脂质分布不对称?
- 膜蛋白如何嵌入、锚定或附着在膜上?
- 膜为什么具有流动性,又为什么不是完全均一的液体?
- 如何研究和重组膜蛋白?
一、细胞膜的五大功能
细胞膜的第一功能是形成边界。脂质双分子层对离子、蛋白质和核酸等亲水分子高度不通透,因此细胞可以把内部环境和外界隔开。
但膜远远不只是边界,它还承担五类功能:
- 区室化:质膜包围细胞,内膜系统把真核细胞内部划分为不同功能区室。
- 门卫作用:转运蛋白选择性地让特定分子进出细胞。
- 信号感知:受体蛋白识别外界信号并启动胞内反应。
- 支架连接:膜蛋白连接细胞外基质、相邻细胞和细胞骨架。
- 能量转化:膜上的转运系统建立离子梯度,驱动 ATP 合成或产生电信号。
所以你理解细胞膜时,不要只记“磷脂双分子层”。更准确地说,膜是一个可流动、可分区、可调控的二维反应平台。
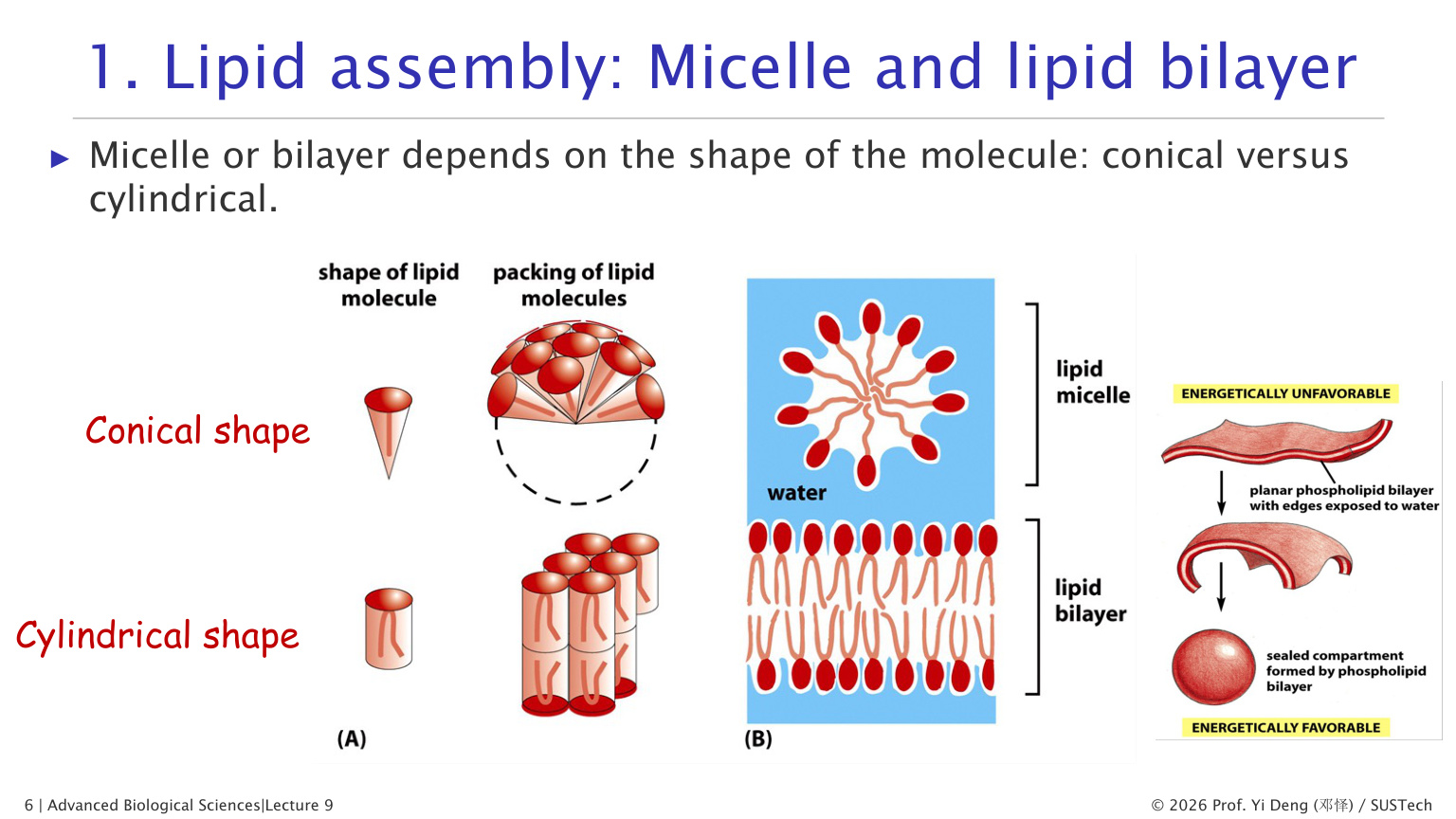
二、脂质为什么会自发形成膜
膜脂是两亲性分子:一端亲水,一端疏水。放入水中后,疏水尾部会尽量避开水,亲水头部朝向水相。
脂质形成什么结构,取决于分子形状:
| 分子形状 | 常见结构 | 直觉理解 |
|---|---|---|
| 圆锥形,头大尾小 | 胶束 micelle | 尾巴聚在内部,头部朝外 |
| 圆柱形,头尾截面积相近 | 脂质双分子层 lipid bilayer | 两层尾部相对,头部朝水 |
双分子层还会自发封闭成囊泡,因为平面边缘会把疏水尾部暴露给水,这在能量上不利。封闭成球形结构后,疏水尾部被埋起来,体系更稳定。
这个物理事实非常重要:细胞膜的形成不需要复杂模板,脂质本身的化学性质就足以驱动膜的自组装。
三、真核细胞膜的三大脂质组分
真核细胞质膜主要由三类脂质组成:
- 磷脂 phospholipids
- 固醇 sterols,动物细胞中主要是胆固醇 cholesterol
- 糖脂 glycolipids
1. 磷脂:膜的主体
磷脂大致分为两类:
- 磷酸甘油酯 phosphoglycerides:以甘油为骨架,是膜中最丰富的磷脂。
- 鞘磷脂 sphingomyelin:以鞘氨醇 sphingosine 为骨架,常富集在质膜外叶。
常见磷酸甘油酯包括:
| 名称 | 缩写 | 头基电荷 | 常见分布 |
|---|---|---|---|
| 磷脂酰胆碱 | PC | 中性 | 主要在外叶 |
| 磷脂酰乙醇胺 | PE | 中性 | 主要在内叶 |
| 磷脂酰丝氨酸 | PS | 负电 | 主要在内叶 |
| 磷脂酰肌醇 | PI | 负电 | 几乎全部在内叶 |
脂肪酸尾部也影响膜性质。饱和尾巴更直,容易紧密排列;不饱和尾巴含顺式双键,会形成弯折,使脂质难以紧密堆积,从而提高膜流动性。
2. 胆固醇:膜流动性的缓冲器
胆固醇具有刚性的四环结构,插在磷脂尾部之间。它对膜流动性的影响具有双向性:
- 低温时,胆固醇阻止脂肪酸尾部紧密排列,防止膜凝固。
- 高温时,胆固醇限制尾部过度运动,降低膜过度流动。
所以胆固醇不是简单地“增加”或“降低”流动性,而是让膜维持在合适的物理状态。
3. 糖脂:细胞表面的识别层
糖脂含有糖基团,在动物细胞中多以鞘脂为基础。它们主要位于质膜外叶,尤其在神经细胞中丰富。
常见类型:
- 脑苷脂 cerebroside:含单个糖基,通常电荷中性。
- 神经节苷脂 ganglioside:含多个糖基,常带有唾液酸,因此带负电。
糖脂的功能包括保护细胞表面、参与细胞识别,并可作为病原体或毒素的受体。例如 GM1 是霍乱毒素的受体。
四、不同膜的脂质组成不同
不同细胞器膜并不是同一种“通用膜”。它们的脂质组成反映各自功能。
例如:
- 细菌膜通常没有胆固醇,PE 含量高。
- 髓磷脂糖脂含量很高,适合神经轴突绝缘。
- 线粒体内膜具有特殊脂质组成,适合能量转化。
- ER 膜是很多脂质合成的起点。
这说明膜脂不仅是结构材料,也参与决定细胞器身份和功能。
五、脂质合成与转运
大多数膜脂在 ER 上合成,包括磷脂、胆固醇和神经酰胺等。
脂质从 ER 到其他膜区室主要有三种路线:
- 囊泡转运:脂质随囊泡出芽和融合移动。
- 膜接触位点:两个细胞器膜靠得很近,脂质可在接触点直接转移。
- 胞质脂质转运蛋白:蛋白从一个膜中提取脂质,再把它送到另一个膜。
注意,膜脂转运并不是随机平均分配。细胞需要把特定脂质富集在特定膜、特定叶层和特定微域中。
六、质膜脂质的不对称性
质膜外叶和内叶组成不同:
- 外叶:糖脂、PC、鞘磷脂较多。
- 内叶:PS、PE、PI 较多。
- 胆固醇:两侧大致都有。
脂质自发从一个叶层翻到另一个叶层非常慢,因为极性头基要穿过疏水核心,能量代价高。细胞依赖三类转位酶维持或改变不对称性:
| 转位酶 | 方向 | 能量 | 功能 |
|---|---|---|---|
| Flippase 翻转酶 | 外叶到内叶 | 需要 ATP | 将 PS/PE 维持在内叶 |
| Floppase 外翻酶 | 内叶到外叶 | 需要 ATP | 将特定脂质转到外叶 |
| Scramblase 扰乱酶 | 双向随机 | 常受 Ca2+ 调控 | 破坏不对称性 |
一个常考例子是凋亡。正常细胞中 PS 位于内叶;凋亡时 scramblase 活化,PS 暴露在外叶,成为巨噬细胞识别的“吃我”信号。
七、磷脂酰肌醇与膜信号
PI 位于质膜内叶,它的肌醇环可以被不同激酶磷酸化,生成多种 PIP 脂质。
重要例子:
- PI(4,5)P2 可被 PLC 切割为 IP3 和 DAG。
- IP3 促进 ER 释放 Ca2+。
- DAG 留在膜上并激活 PKC。
- PI(3,4,5)P3 可招募带 PH 结构域的信号蛋白,如 Akt。
- PTEN 可去磷酸化 PI(3,4,5)P3,是重要肿瘤抑制因子。
所以 PI 类脂质虽然含量不高,却是膜上信号组织的关键坐标。
八、脂滴:储存脂质的特殊细胞器
脂滴 lipid droplet 由磷脂单分子层包裹中性脂质核心,核心主要含三酰甘油 TAG 和胆固醇酯 CE。
它们从 ER 形成,功能包括:
- 储存能量。
- 缓冲脂质毒性。
- 维持脂质稳态。
- 为膜合成和代谢提供脂质来源。
脂滴不是“油滴垃圾堆”,而是可调控的代谢细胞器。
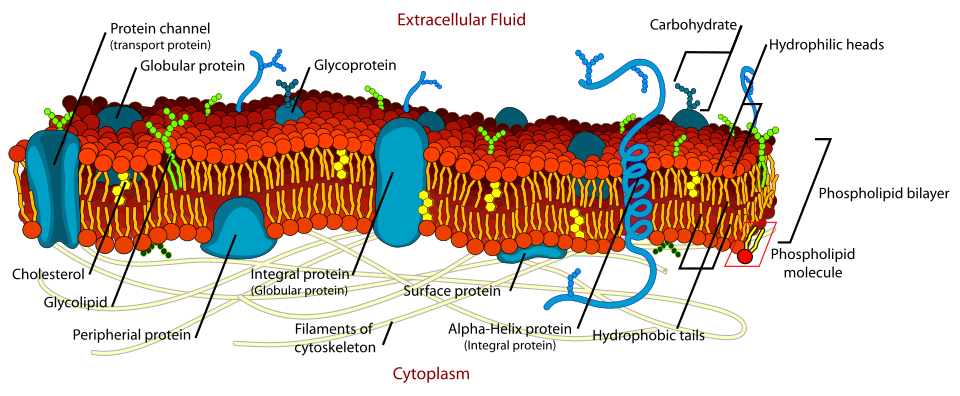
九、膜蛋白的功能
膜的很多关键功能由膜蛋白执行,包括:
- 转运:离子通道、转运体、泵。
- 受体:识别激素、生长因子、神经递质等。
- 酶:把反应限制在膜表面或特定细胞器上。
- 连接:把细胞外基质或相邻细胞连接到细胞骨架。
- 能量转换:如电子传递链和 ATP 合酶。
- 膜变形:参与囊泡形成、膜融合和细胞形态变化。
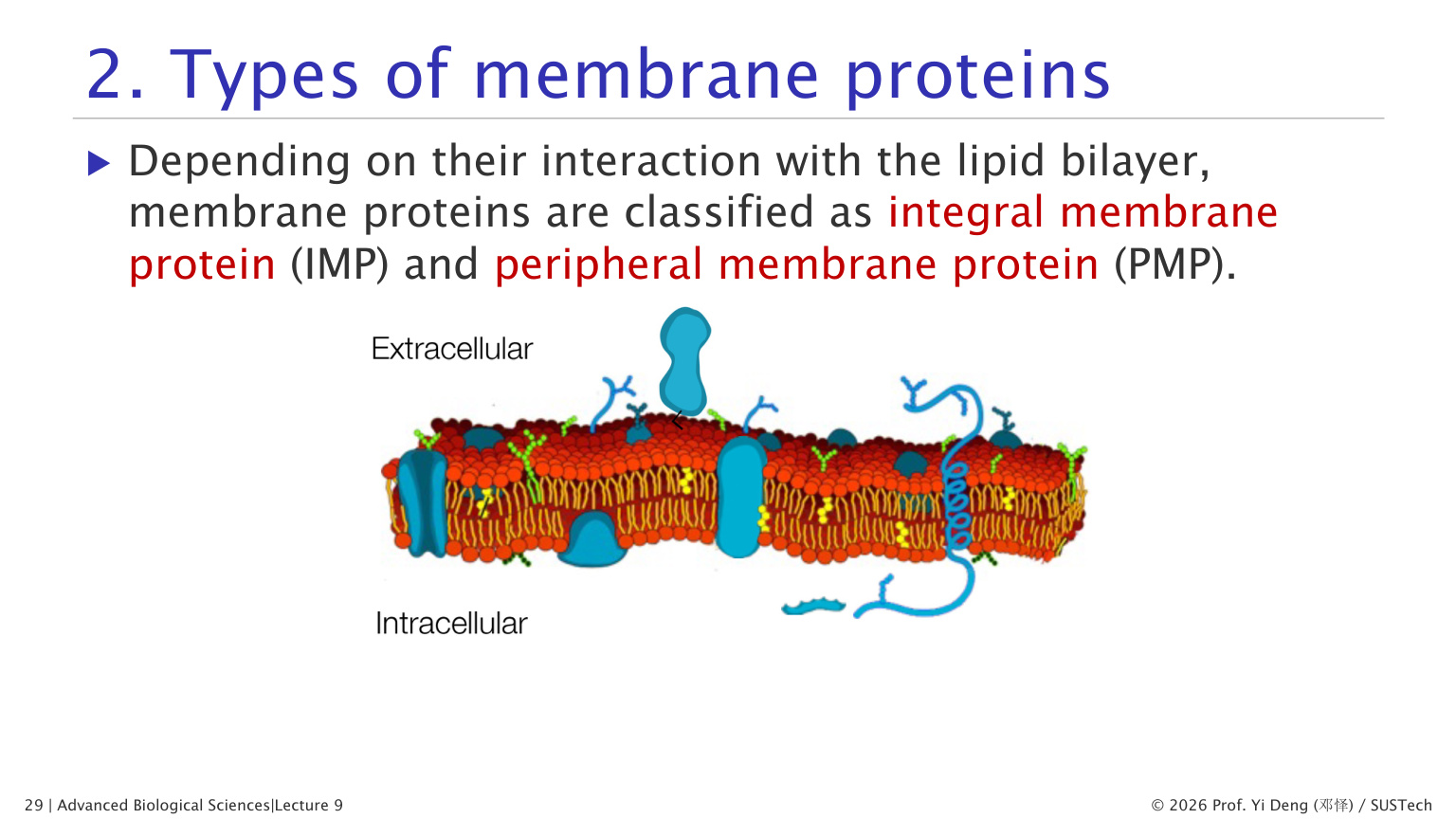
十、膜蛋白的类型
1. 内在膜蛋白 integral membrane proteins
内在膜蛋白与脂双层结合紧密,通常需要去污剂才能提取。
常见形式:
- 跨膜蛋白:一段或多段跨过整个脂双层。
- 单层嵌入蛋白:只嵌入一个叶层。
- 脂质锚定蛋白:通过共价连接的脂质锚定位于膜上。
脂质锚包括:
- Myristoyl anchor,常连在 N 端甘氨酸。
- Palmitoyl anchor,常连在半胱氨酸。
- Prenyl anchor,常见于 Ras、Rab 等小 G 蛋白。
- GPI anchor,位于膜外叶,常见于细胞表面蛋白。
2. 外周膜蛋白 peripheral membrane proteins
外周膜蛋白不插入疏水核心,而是通过非共价相互作用结合在膜表面,常可用高盐或改变 pH 洗脱。
十一、跨膜蛋白的结构特征
多数跨膜蛋白用 alpha 螺旋穿过脂双层。一个跨膜 alpha 螺旋通常约 20-30 个氨基酸,富含疏水残基。
跨膜区可以用疏水性图 hydropathy plot 预测:连续疏水残基会形成明显峰值。
另一类跨膜结构是 beta 桶,常见于:
- 革兰氏阴性菌外膜。
- 线粒体外膜。
- 叶绿体外膜。
beta 桶可以形成孔蛋白、受体或转运相关结构。
十二、糖被与细胞识别
细胞表面常覆盖糖链,形成糖被 glycocalyx。糖链来自糖蛋白和糖脂,并且总是位于细胞外侧或细胞器腔面。
糖被的功能:
- 保护细胞表面。
- 增加润滑性。
- 参与细胞识别。
- 介导细胞黏附和免疫识别。
Lectins 是糖结合蛋白,能特异识别糖链。比如血管内皮和免疫细胞之间的识别就常依赖糖链和 lectin。
十三、膜的流动性
膜不是刚性壳,而是流体。脂质可以:
- 侧向扩散 lateral diffusion。
- 旋转 rotation。
- 弯曲 flexion。
- 很慢地跨叶层翻转 flip-flop。
影响流动性的因素:
- 不饱和脂肪酸越多,流动性越高。
- 脂肪酸链越短,流动性越高。
- 温度升高,流动性升高。
- 胆固醇起缓冲作用。
低温时膜可进入凝胶相,高温时进入液晶相。细胞需要把膜维持在合适的流动范围内,太硬或太软都不利于功能。
十四、脂质筏与膜微域
质膜并不是完全均一的二维液体。某些区域富含胆固醇、鞘磷脂和糖脂,形成更厚、更有序的微域,常称为脂质筏 lipid rafts。
脂质筏可富集:
- GPI 锚定蛋白。
- 某些跨膜受体。
- Src 家族激酶等信号蛋白。
其功能是把相关分子集中在同一区域,提高信号传递和膜组织效率。
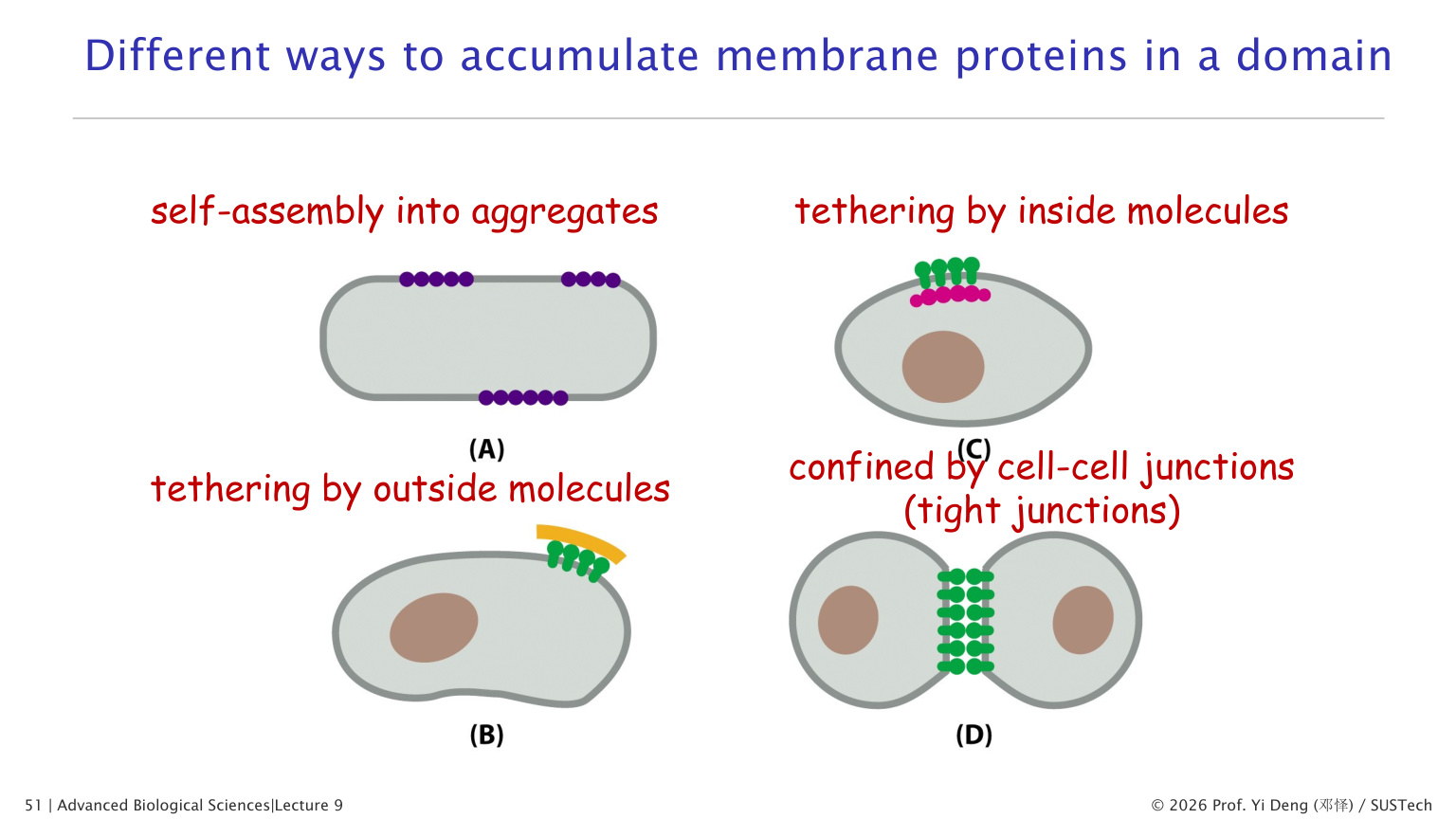
十五、膜蛋白移动为什么会受限制
膜蛋白可以侧向扩散,但很多蛋白并不完全自由。
限制机制包括:
- 蛋白自组装成大复合物。
- 与细胞外基质结合。
- 与细胞骨架结合。
- 被紧密连接等细胞连接限制在特定膜域。
- 被皮层细胞骨架形成的“围栏”限制。
红细胞中的 spectrin 网络就是经典例子。Spectrin、ankyrin、actin、band 3 等形成膜骨架,维持红细胞形态并限制膜蛋白扩散。
十六、研究膜流动性和膜蛋白的方法
1. FRAP
FRAP 是 fluorescence recovery after photobleaching。实验思路是用荧光标记膜分子,用强光漂白一小块区域,然后观察周围未漂白分子扩散进入该区域的速度。
它可以反映:
- 分子在膜平面内的扩散速度。
- 可移动分子的比例。
2. 细胞融合实验
把人细胞和小鼠细胞融合,并用不同颜色标记膜蛋白。融合后初期两类膜蛋白分开,随后逐渐混合,说明膜蛋白可以在膜平面内侧向扩散。
3. 单粒子追踪
高速单粒子追踪可以看到膜蛋白不是简单自由扩散,而常在小范围内受限扩散,偶尔跳到相邻区域。
十七、去污剂与膜蛋白重组
去污剂 detergent 是研究膜蛋白的重要工具。它们也是两亲性分子,能把膜脂溶解成混合胶束。
常见类型:
| 类型 | 代表 | 特点 |
|---|---|---|
| 离子型 | SDS | 溶解力强,但会使蛋白变性 |
| 非离子型 | Triton X-100、NP-40、beta-octyl glucoside | 较温和,可保留部分蛋白活性 |
研究膜蛋白常需要功能重组:
1. 用去污剂溶解膜。 2. 纯化目标膜蛋白。 3. 加入磷脂。 4. 去除去污剂。 5. 形成含功能蛋白的蛋白脂质体 proteoliposome。
纳米盘 nanodisc
Nanodisc 是一小片脂质双分子层,边缘由蛋白“腰带”包围,使其可溶于水。膜蛋白插入 nanodisc 后可更接近天然状态,因此常用于结构生物学,尤其是冷冻电镜研究。
十八、常见误区
- 误区一:膜只是“脂质袋子”。实际上膜蛋白和糖链决定了大量功能。
- 误区二:所有脂质都只是结构材料。PI、PIP、DAG、鞘脂代谢物都可作为信号分子。
- 误区三:膜两侧脂质一样。质膜叶层不对称是重要功能基础。
- 误区四:膜越流动越好。流动性必须维持在合适范围。
- 误区五:膜蛋白都能自由移动。细胞骨架、细胞连接和微域会限制扩散。
十九、你必须会的关键词
- amphipathic molecule
- lipid bilayer
- phosphoglyceride
- sphingomyelin
- cholesterol
- glycolipid
- lipid asymmetry
- flippase
- floppase
- scramblase
- phosphoinositide
- lipid droplet
- integral membrane protein
- peripheral membrane protein
- GPI anchor
- glycocalyx
- membrane fluidity
- lipid raft
- FRAP
- detergent
- nanodisc
二十、自测题
1. 为什么双尾磷脂更容易形成双分子层,而不是胶束? 2. 胆固醇如何在低温和高温下分别影响膜流动性? 3. 为什么 PS 暴露到质膜外叶可以作为凋亡信号? 4. PI(4,5)P2 和 PI(3,4,5)P3 各自参与哪些信号事件? 5. 内在膜蛋白和外周膜蛋白如何区分? 6. alpha 螺旋跨膜区为什么富含疏水氨基酸? 7. FRAP 实验能告诉我们什么,不能告诉我们什么? 8. 为什么研究膜蛋白需要去污剂或 nanodisc?
二十一、考前速记版
膜的本质是两亲性脂质自组装成的双分子层。真核质膜主要含磷脂、胆固醇和糖脂;两侧脂质分布不对称,靠 flippase、floppase 和 scramblase 调控。膜蛋白有跨膜、单层嵌入、脂质锚定和外周结合等形式,是膜功能的主要执行者。膜具有流动性,但流动性受脂肪酸组成、温度、胆固醇和细胞骨架限制。脂质筏可富集信号分子。研究膜蛋白常用去污剂、蛋白脂质体和 nanodisc。