NLR 与抗病小体
从细胞内受体到钙离子通道——植物如何用"自杀式攻击"阻止病原体入侵
4.1引言
当病原体成功突破植物表面的 PTI (Pattern-Triggered Immunity) 防线后,它们分泌的效应子 (effector) 进入植物细胞内部,试图抑制免疫信号并劫持宿主代谢。面对这一威胁,植物演化出一类细胞内免疫受体——NLR 蛋白 (Nucleotide-binding Leucine-rich Repeat),它们能够直接或间接识别效应子的活动,并触发强烈的防御响应。
NLR 介导的免疫通常伴随过敏性坏死反应 (Hypersensitive Response, HR)——受感染细胞主动死亡,将病原体封锁在坏死区域内。长期以来,NLR 激活与 HR 执行之间的分子联系一直是植物免疫学的核心谜题。直到 2019 年,ZAR1 抗病小体 (resistosome) 结构的解析,才从原子层面揭示了 NLR 蛋白从信号感知到细胞死亡执行的完整机制——NLR 本身即是执行细胞死亡的钙离子通道。这一发现从根本上改写了对 ETI 信号转导的理解。
4.2NLR 蛋白的分类与结构域组织
植物 NLR 蛋白属于 STAND 超家族 (Signal Transduction ATPases with Numerous Domains),与动物先天免疫中的 NLR 和 inflammasome 具有进化上的同源性。根据 N 端效应结构域的差异,植物 NLR 分为三大类:
| 类型 | N 端结构域 | 代表成员 | 功能特征 |
|---|---|---|---|
| CNL | CC (Coiled-Coil) | ZAR1, RPM1, Rx | 激活后形成五聚体,CC 结构域直接插膜形成钙离子通道 |
| TNL | TIR (Toll/Interleukin-1 Receptor) | RPP1, ROQ1, SNC1 | 激活后 TIR 结构域获得 NADase 酶活性,通过 EDS1 传递信号 |
| RNL | RPW8 (CCR) | NRG1, ADR1 | 作为 helper NLR,在 TNL 下游执行细胞死亡 |
三类 NLR 均共享保守的中央 NB-ARC 结构域 (Nucleotide-Binding domain shared with Apaf-1, R proteins, and CED-4) 和 C 端 LRR 结构域 (Leucine-Rich Repeat)。NB-ARC 结构域是分子开关的核心:静息状态下结合 ADP,维持蛋白的自抑制构象;激活后 ADP 被交换为 ATP,触发构象变化并驱动寡聚化。
4.3NLR 的激活机制:从自抑制到寡聚化
NLR 在未被激活时以自抑制构象 (autoinhibited conformation) 存在。LRR 结构域折叠回来包裹 NB-ARC 结构域,阻止其核苷酸交换和寡聚化。这种精密的自抑制机制确保 NLR 不会在没有病原体的情况下被意外激活——因为 NLR 的激活通常意味着细胞死亡,误激活将对植物自身造成严重损害。
当效应子被识别后(无论是直接结合还是通过感知宿主蛋白的修饰),LRR 结构域的构象发生改变,释放对 NB-ARC 的抑制。随后发生以下关键事件:
- 核苷酸交换:NB-ARC 结构域释放 ADP,结合 ATP。这是激活的关键分子开关。
- 构象开放:ATP 结合诱导 NB-ARC 结构域的大幅构象变化,暴露出此前被掩埋的寡聚化界面。
- 寡聚化组装:多个激活态 NLR 单体通过 NB-ARC 结构域的相互作用组装成轮状寡聚体。CNL 通常形成五聚体,TNL 形成四聚体。
- N 端效应结构域暴露:寡聚化将 N 端 CC 或 TIR 结构域聚集在一起,执行下游功能——CNL 的 CC 结构域直接形成膜孔,TNL 的 TIR 结构域获得 NADase 活性。
4.4ZAR1 抗病小体——里程碑式的结构突破
2019 年,柴继杰团队与周俭民团队合作,在 Science 杂志连发两篇论文,报道了拟南芥 ZAR1 蛋白在静息态、中间态和激活态三种构象下的冷冻电镜结构 (Wang et al., 2019a, b)。这是首个完整的植物 NLR 激活态结构,也标志着抗病小体 (resistosome) 概念的正式确立。
图 4.1 ZAR1 抗病小体的组装过程。 (A) 静息态:ZAR1LRR-RKS1 二元复合体,NB-ARC 结合 ADP,蛋白处于自抑制构象。 (B) 中间态:效应子 AvrAC 修饰 PBL2 后,PBL2UMP 被 RKS1 捕获,触发 ZAR1 的 ADP→ATP 交换。 (C) 激活态:五个 ZAR1 单体组装为轮状五聚体,N 端 CC 结构域形成漏斗状结构,插入质膜形成钙离子通道。 改编自 Wang et al., 2019, Science.
ZAR1 的激活过程清晰地展示了从效应子识别到细胞死亡的完整分子逻辑:
- 效应子活动的间接感知:丁香假单胞菌 (Pseudomonas syringae) 的 III 型效应子 AvrAC 将 UMP 基团转移到宿主激酶 PBL2 上,干扰其在 PTI 中的信号功能。
- 诱饵蛋白的捕获:被修饰的 PBL2UMP 被 ZAR1 复合体中的假激酶 RKS1 识别和结合。PBL2 在此充当"诱饵" (decoy),其被效应子修饰本身就是危险信号。
- 核苷酸交换与寡聚化:PBL2UMP 的结合触发 ZAR1 的 ADP→dATP/ATP 交换,诱导构象开放,五个 ZAR1-RKS1-PBL2UMP 三元复合体组装为轮状五聚体。
- 膜孔形成:五聚体的 N 端 CC (α1 螺旋) 结构域聚集形成漏斗状结构,插入质膜,形成选择性钙离子通道。
- 钙内流与细胞死亡:胞外 Ca²⁺ 大量内流,触发下游 HR 相关的信号通路,最终导致细胞程序性死亡。
在 ZAR1 抗病小体结构解析之前,领域对 NLR 激活后的信号传递机制存在长期争论。主流假设认为 NLR 激活后通过招募下游信号蛋白(类似动物 NLR 招募 caspase 的模式)间接启动防御。ZAR1 结构的揭示推翻了这一假设,证明植物 CNL 类 NLR 本身即是效应器 (executor)——它直接在质膜上打孔,而非通过中间信号分子传递信息。这一概念转变重新定义了植物 ETI 的信号转导范式,并将植物 NLR 与动物免疫中的 gasdermin 成孔蛋白进行了功能类比,揭示了跨界免疫的深层统一性。
4.5TNL 的信号转导:NADase 活性与 EDS1 枢纽
与 CNL 直接成孔不同,TNL 类 NLR 的激活后信号传递依赖一条截然不同的通路。TNL 激活后组装为四聚体抗病小体,其 TIR 结构域在寡聚化后获得 NADase 酶活性,将 NAD⁺ 水解为一系列小分子信号产物。
这些小分子信号(包括变体环化 ADP-核糖, v-cADPR 等)被脂酶蛋白 EDS1 (Enhanced Disease Susceptibility 1) 感知。EDS1 与其异源二聚体伙伴 PAD4 或 SAG101 形成复合体后,分别激活下游的 helper NLR——ADR1 或 NRG1。最终,这些 RNL 类 helper NLR 执行细胞死亡和免疫信号输出。
TIR 结构域的 NADase 酶活性是 2019 年前后由多个团队独立发现的 (Horsefield et al., 2019; Wan et al., 2019)。此前,TIR 结构域被认为仅作为蛋白-蛋白互作的支架结构域发挥功能。酶活性的发现揭示了 TNL 信号转导的全新生化机制,也解释了为什么 TNL 和 CNL 的下游通路如此不同——CNL 自身成孔,而 TNL 需要通过酶产物传递信号给 helper NLR 来执行死亡。
4.6Integrated Decoy 与感知多样性的扩展
NLR 如何识别效应蛋白?最初的"基因对基因"假说暗示直接的受体-配体结合,但保卫假说已经表明间接识别可能更为普遍 → 第1章 1.3节。"整合诱饵"(Integrated Decoy / Integrated Domain, ID)模型将这一概念推向了新的高度。
从保卫到整合:概念的进化
保卫假说(Guard Model)认为 NLR 监测效应蛋白的靶标蛋白——当靶标被修饰时,NLR 被激活。但如果被监测的蛋白逐渐丧失其原始功能,专门演化为"诱饵"(Decoy),我们就得到了诱饵模型 (van der Hoorn & Kamoun, 2008)。整合诱饵模型更进一步:将诱饵结构域直接整合到 NLR 蛋白自身中,形成一个"受体-诱饵"的融合蛋白 (Cesari et al., 2014)。
具体而言,许多 NLR 蛋白在经典的 NB-ARC-LRR 结构之外,还包含一个"非典型"结构域——即整合域(ID)。这些 ID 往往是效应蛋白在宿主中的真实靶标的同源序列。例如:
- 水稻 RGA5:其 C 端整合了一个重金属相关域(HMA),是M. oryzae效应蛋白 AVR-Pia 的直接结合靶标。AVR-Pia 结合 HMA 域后,RGA5 与其配对 NLR RGA4 协同激活免疫 (Cesari et al., 2013)。
- 拟南芥 RRS1:整合了一个 WRKY 转录因子域。Ralstonia solanacearum的效应蛋白 PopP2 是一种乙酰转移酶,靶向 WRKY 蛋白进行乙酰化修饰以抑制免疫。当 PopP2 修饰 RRS1 中的 WRKY 域时,RRS1 与配对 NLR RPS4 协同激活 ETI (Le Roux et al., 2015; Sarris et al., 2015)。
- 水稻 Pik-1:整合了一个 HMA 域,直接结合M. oryzae效应蛋白 AVR-PikD,与配对 NLR Pik-2 协同工作 (Maqbool et al., 2015)。
图 4.2 整合诱饵与 sensor-helper 分工。携带整合域的 sensor NLR 负责捕获效应蛋白活动,helper NLR 负责稳定输出。识别端可以快速进化,输出端保持保守,这使 NLR 网络既能追踪病原变化,又能维持可靠的信号执行。
NLR 配对:sensor-executor 的最小单元
一个引人注目的规律是:携带 ID 的 NLR 几乎总是成对出现——一个 sensor NLR(携带 ID,负责识别)和一个 executor/helper NLR(负责信号输出),两者基因在基因组上通常头对头或尾对尾排列 (Cesari et al., 2014)。这种配对架构赋予了极大的进化灵活性:sensor NLR 的 ID 可以快速变异以追踪效应蛋白的进化,而 executor NLR 保持保守以维持可靠的信号输出。
生物信息学分析显示,植物 NLR 组中整合的域类型极为多样——包括激酶域、WRKY 域、HMA 域、BED 锌指域等,总计超过50种不同的整合域类型 (Kroj et al., 2016; Sarris et al., 2016)。这些整合域的多样性可能反映了效应蛋白靶标的多样性,暗示 ID-NLR 是植物应对效应蛋白多样化的一种高效进化策略。
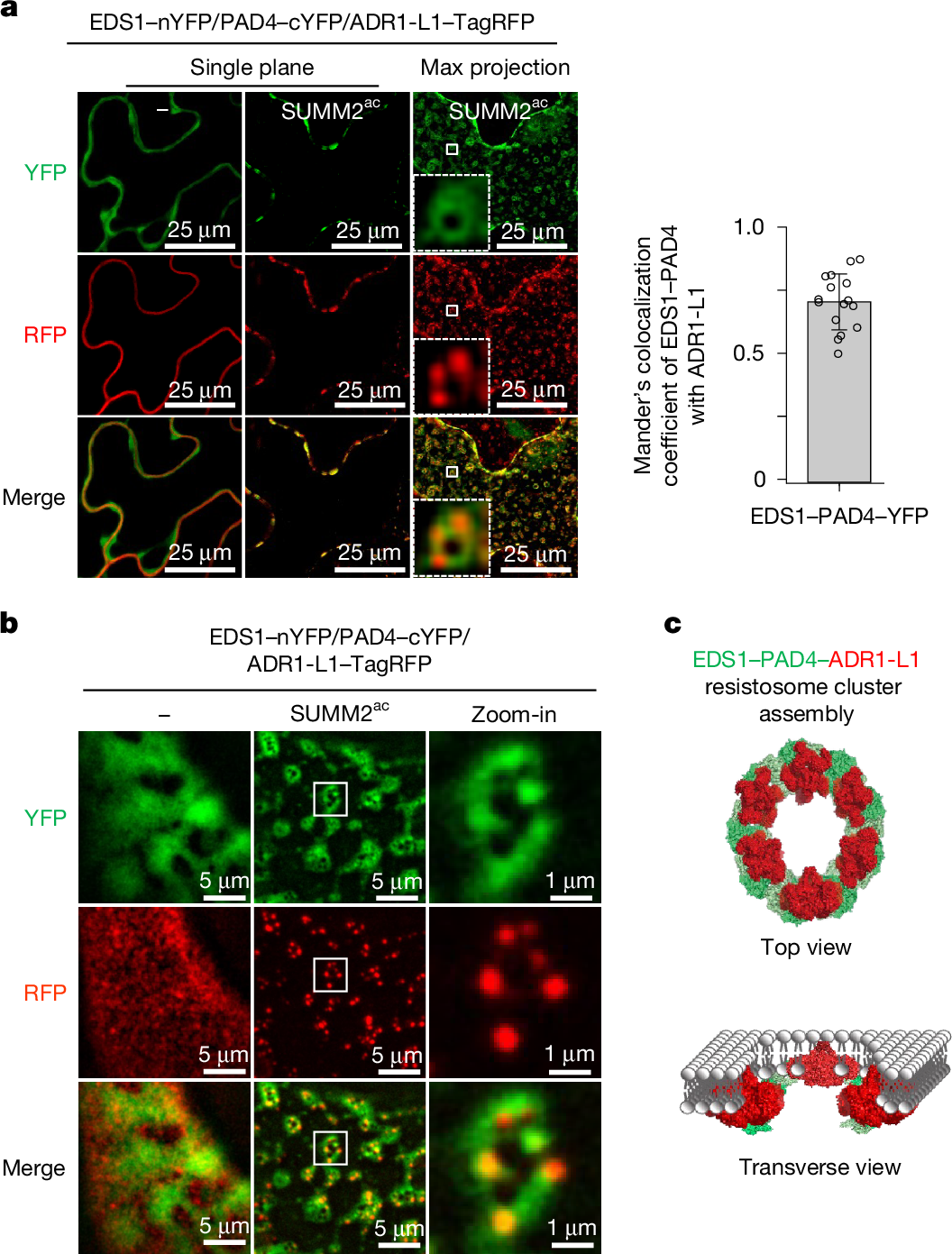
图 4.P1 论文原图:helper NLR 抗病小体簇把 NLR 输出从线性通路推向高阶组装。原图为 Ge et al. (2026) Nature Fig. 4,DOI: 10.1038/s41586-026-10215-1。该图展示 SUMM2 激活后 EDS1-PAD4 与 ADR1-L1 的空间组织和簇状组装,直接对应本章关于 sensor-helper 分工和 NLR 输出阈值的讨论。依据 CC BY-NC-ND 4.0 原样复用;图片未作裁剪、改色或内容改动。
整合诱饵模型揭示了植物免疫的一个深刻的进化智慧:把效应蛋白的攻击靶标变成识别陷阱。效应蛋白进化来修饰宿主蛋白 X → 植物把 X 的结构域整合进 NLR → 效应蛋白"上钩"修饰整合域 → NLR 激活。这种策略的精妙之处在于,效应蛋白越是精准地攻击其靶标,就越容易被"整合了靶标的 NLR"所识别。这为合理设计新型抗病 NLR 提供了明确的工程思路 → 第14章。
4.7NLR 网络:sensor 与 helper 的分工协作
随着越来越多 NLR 的功能被揭示,领域认识到植物体内的 NLR 并非各自为战,而是组成了一个功能网络。根据功能分工,NLR 可分为:
Sensor NLR:负责识别特定的效应子或效应子活动。它们的 LRR 或整合结构域 (Integrated Domain, ID) 提供识别特异性,但自身可能不直接执行免疫信号输出。
Helper NLR:接收 sensor NLR 的信号,执行下游功能(如成孔、激活防御基因)。典型的 helper NLR 包括 NRG1、ADR1(RNL 类)和 NRC 家族(CNL 类)。
这种 sensor-helper 的分工模式具有重要的进化意义:sensor NLR 可以快速多样化以应对不断变化的效应子,而 helper NLR 保持相对保守,维持可靠的信号输出。这类似于适应性免疫中"识别"与"效应"功能的分离,是进化效率的体现。
2026 年的结构研究进一步把 helper NLR 从“下游执行器”推进到“可聚合的信号平台”。Ge et al. (2026) 发现,CNL 型 sensor NLR SUMM2 激活后,可以诱导 EDS1-PAD4 与 ADR1-L1 组装成 helper NLR 抗病小体簇,而不是一条简单的线性通路。这个结果提示:helper NLR 的寡聚状态可能决定输出阈值、细胞死亡强度和防御基因激活比例,是未来解耦“有效防御”与“组织损伤”的关键调控层。
4.8过敏性坏死反应 (HR) 的分子基础
HR 是 ETI 最显著的表型输出——受感染的细胞及其周围少量细胞迅速死亡,在叶片上形成可见的坏死斑点,将病原体(尤其是活体营养型病原体)封锁在死亡组织中。HR 的核心分子执行者现在被认为是 NLR 抗病小体形成的膜孔本身,以及其导致的 Ca²⁺ 内流。
值得注意的是,细胞死亡与免疫信号是否可以解耦 (uncoupling),是领域内的重要争论之一。部分证据表明,NLR 可以在不引发明显 HR 的情况下激活免疫基因表达和抗病性,提示 NLR 的功能输出可能比"全有或全无"的细胞死亡更为精细。
NLR 介导的细胞死亡与免疫信号输出能否被彻底解耦?
如果可以解耦——即保留 NLR 的免疫信号输出(防御基因激活、SA 积累等)同时消除细胞死亡——那将在育种上具有巨大价值:抗性增强但没有 HR 带来的组织损伤代价。初步证据暗示 CNL 的成孔和信号输出可能有部分独立性(低水平的 Ca²⁺ 内流足以激活信号,但不足以杀死细胞),但这一假说的普遍适用性尚待验证。对 TNL 来说,由于信号通过酶产物传递给 helper NLR,理论上可以通过工程化 helper NLR 的活性来调控输出水平 → 第14章。
4.9里程碑研究思路拆解
里程碑 1:Wang et al. (2019a, 2019b) — ZAR1 抗病小体的结构解析
面对的问题:NLR 蛋白激活后如何触发细胞死亡?尽管遗传学证据已积累数十年,但缺乏直接的结构和生化证据解释 NLR 从"感知效应子"到"执行死亡"的跨越。
关键思路:团队选择 ZAR1 作为研究对象,因为其生化性质较好且已有较完善的遗传背景。核心策略是在体外重建完整的激活复合体,然后用冷冻电镜逐步解析从静息态到激活态的构象变化。这一思路的关键创新在于:不是只解析一个终态结构,而是捕捉了三个连续状态(静息态、中间态、激活态),使得整个激活过程的动态机制一览无遗。
关键证据链:
① 冷冻电镜结构显示 ZAR1 激活后形成五聚体,CC 结构域组成漏斗状结构 → ② 结构预测该漏斗可插入脂质膜 → ③ 脂质体实验验证 ZAR1 五聚体确实在膜上成孔 → ④ 电生理实验证明该孔道具有钙离子选择性 → ⑤ 植物体内实验证实 Ca²⁺ 内流对于 HR 是必需的。
影响:该工作将 NLR 从"信号感受器"重新定义为"信号执行器",彻底改变了对植物 ETI 分子机制的理解,并被迅速写入教科书。
里程碑 2:Horsefield et al. (2019) & Wan et al. (2019) — TIR 域的 NADase 活性
面对的问题:TNL 类 NLR 的 TIR 结构域如何传递信号?此前认为 TIR 仅作蛋白互作支架,但遗传证据表明其自身的某种酶活性对信号传递是必需的。
关键思路:两个独立团队通过结构和生化分析发现,TIR 域在寡聚化后获得 NAD⁺ 水解酶(NADase)活性,将 NAD⁺ 切割为烟酰胺和变体环化 ADP-核糖(v-cADPR)等产物。
关键证据链:(1)纯化 TIR 域体外水解 NAD⁺;(2)催化位点关键残基突变(如 E/A 突变)消除酶活和体内免疫功能;(3)产物 v-cADPR 被 EDS1 复合体感知。
影响:揭示了 TNL 信号转导的全新生化机制,解释了 TNL 和 CNL 下游通路分歧的分子基础。也建立了植物 TIR 与动物 SARM1(一种同样具有 NADase 活性的 TIR 域蛋白)之间的功能平行关系。
里程碑 3:Cesari et al. (2014) & Le Roux et al. (2015) — 整合诱饵模型
面对的问题:NLR 如何应对效应蛋白靶标的多样性?如果效应蛋白靶向不同的宿主蛋白,植物是否需要为每个靶标单独演化一个 NLR?
关键思路:通过比较基因组学分析发现,许多 NLR 在经典结构域之外整合了额外的"非典型域"(ID),这些 ID 的序列与已知效应蛋白靶标同源。由此提出:NLR 通过将效应蛋白靶标整合为自身的一部分,以"钓鱼"方式捕获效应蛋白。
关键证据链:(1)水稻 RGA5 的 HMA 域直接结合 AVR-Pia;(2)拟南芥 RRS1 的 WRKY 域被 PopP2 乙酰化后触发 RPS4 激活;(3)全基因组分析揭示超过50种不同的 ID 类型。
影响:为合理设计新型 NLR 提供了清晰的策略框架:鉴定效应蛋白靶标 → 将靶标域工程整合进 NLR sensor → 配对 executor NLR。这一策略已在水稻抗稻瘟病中初步验证。
4.10当前争论与未解问题
- 抗病小体的通道选择性和调控机制尚不清楚。ZAR1 孔道对 Ca²⁺ 的选择性如何实现?是否存在内源性调控因子控制孔道的开闭?
- NLR 是否存在非细胞死亡依赖的免疫输出?部分实验表明 NLR 可以在不引发 HR 的情况下激活转录重编程,但分子机制不明。如何解耦"死亡"与"免疫"?
- 不同 NLR 的寡聚化是否遵循统一机制?CNL 通常形成五聚体,TNL 形成四聚体,但是否存在其他化学计量比?寡聚数目是否影响功能输出的强度和类型?
- NLR 的亚细胞定位如何影响其功能?部分 NLR 在激活前定位于细胞核,激活后是否需要重新定位到质膜才能成孔?核定位的 NLR 执行什么功能?
- NLR 网络的涌现性质。多个 sensor NLR 是否可以同时激活同一个 helper NLR?信号如何整合?是否存在阈值效应?
2025 年的可激活 NLR 工程研究进一步把这个问题推向应用层面。Wang et al. (2025) 将天然自激活 NLR 改造成“病原蛋白酶激活”的免疫开关:只有当目标病原的特异性蛋白酶切割设计好的连接肽时,NLR 才解除抑制并触发广谱免疫。这一策略的重要性不只在于获得抗性,而在于它提供了一个新的设计原则——不是直接改变 NLR 的效应子识别界面,而是把病原体的保守酶活转化为免疫触发信号。结合 Ge et al. (2026) 对 helper NLR 抗病小体簇的解析,未来工程化的关键不只是“让 NLR 开启”,而是控制它开启后的聚合状态和输出强度。
4.11关键实验方法
| 实验方法 | 用途 | 经典文献 |
|---|---|---|
| 冷冻电镜 (Cryo-EM) | 解析 NLR/抗病小体的三维结构 | Wang et al., 2019, Science |
| 电解质渗漏 (Electrolyte Leakage) | 定量检测 HR 细胞死亡程度 | 标准免疫表型方法 |
| 脂质体重建 (Liposome Reconstitution) | 体外验证 NLR 的成孔活性和离子选择性 | Bi et al., 2021, Nature |
| Co-IP / 免疫共沉淀 | 验证 NLR 复合体的蛋白互作 | 广泛使用 |
| 体外 NADase 活性测定 | 检测 TNL 的 TIR 结构域酶活性 | Horsefield et al., 2019, Science |
| 自激活突变体 (Autoactive mutant) | 模拟 NLR 持续激活状态,验证功能 | 广泛使用 |
4.12推荐阅读
🔴 必读
🟡 重要
🟢 拓展
4.13参考文献
- Adachi H, Derevnina L, Kamoun S. NLR singletons, pairs, and networks: evolution, assembly, and regulation of the intracellular immunoreceptor circuitry of plants. Curr Opin Plant Biol, 2019, 50: 121–131.
- Bi G, Su M, Li N, et al. The ZAR1 resistosome is a calcium-permeable channel triggering plant immune signaling. Cell, 2021, 184: 3528–3541.
- Cesari S, Thouri M, Broz P, et al. The rice resistance protein pair RGA4/RGA5 recognizes the Magnaporthe oryzae effectors AVR-Pia and AVR1-CO39 by direct binding. Plant Cell, 2013, 25: 1463–1481.
- Cesari S, Bernoux M, Moncuquet P, et al. A novel conserved mechanism for plant NLR protein pairs: the "integrated decoy" hypothesis. Front Plant Sci, 2014, 5: 606.
- Dongus JA, Parker JE. EDS1 signalling: at the nexus of intracellular and surface receptor immunity. Curr Opin Plant Biol, 2021, 62: 102039.
- Ge D, Ortiz-Morea FA, Xie Y, et al. Assembly of helper NLR resistosome clusters upon activation of a coiled-coil NLR. Nature, 2026, 652: 251–258. DOI: 10.1038/s41586-026-10215-1.
- Horsefield S, Burdett H, Zhang X, et al. NAD⁺ cleavage activity by animal and plant TIR domains in cell death pathways. Science, 2019, 365: 793–799.
- Kourelis J, van der Hoorn RAL. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell, 2018, 30: 285–299.
- Kroj T, Chanclud E, Michel-Romiti C, et al. Integration of decoy domains derived from protein targets of pathogen effectors into plant immune receptors is widespread. New Phytol, 2016, 210: 618–626.
- Lapin D, Bhandari DD, Parker JE. Origins and immunity networking functions of EDS1 family proteins. Annu Rev Phytopathol, 2022, 60: 253–276.
- Le Roux C, Huet G, Jauneau A, et al. A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity. Cell, 2015, 161: 1074–1088.
- Maqbool A, Saitoh H, Franceschetti M, et al. Structural basis of pathogen recognition by an integrated HMA domain in a plant NLR immune receptor. eLife, 2015, 4: e08709.
- Sarris PF, Duxbury Z, Huh SU, et al. A plant immune receptor detects pathogen effectors that target WRKY transcription factors. Cell, 2015, 161: 1089–1100.
- Sarris PF, Cevik V, Dagdas G, et al. Comparative analysis of plant immune receptor architectures uncovers host proteins likely targeted by pathogens. BMC Biol, 2016, 14: 8.
- van der Hoorn RAL, Kamoun S. From guard to decoy: a new model for perception of plant pathogen effectors. Plant Cell, 2008, 20: 2009–2017.
- Wan L, Essuman K, Anderson RG, et al. TIR domains of plant immune receptors are NAD⁺-cleaving enzymes that promote cell death. Science, 2019, 365: 799–803.
- Wang X, Ji C, Wang L, et al. Remodelling autoactive NLRs for broad-spectrum immunity in plants. Nature, 2025, 645: 737–745. DOI: 10.1038/s41586-025-09252-z.
- Wang J, Hu M, Wang J, et al. Reconstitution and structure of a plant NLR resistosome conferring immunity. Science, 2019a, 364: eaav5870.
- Wang J, Wang J, Hu M, et al. Ligand-triggered allosteric ADP release primes a plant NLR complex. Science, 2019b, 364: eaav5868.